Parallel bedding in a Paleocene turbidite succession, Point San Pedro, California. The thickness of individual beds varies little along their lateral extent, at least within the confines of the outcrop; our view of bedding planes is limited to their 2D extent. The thickest bed is about 50 cm. Geologist’s shoe on the bottom right.
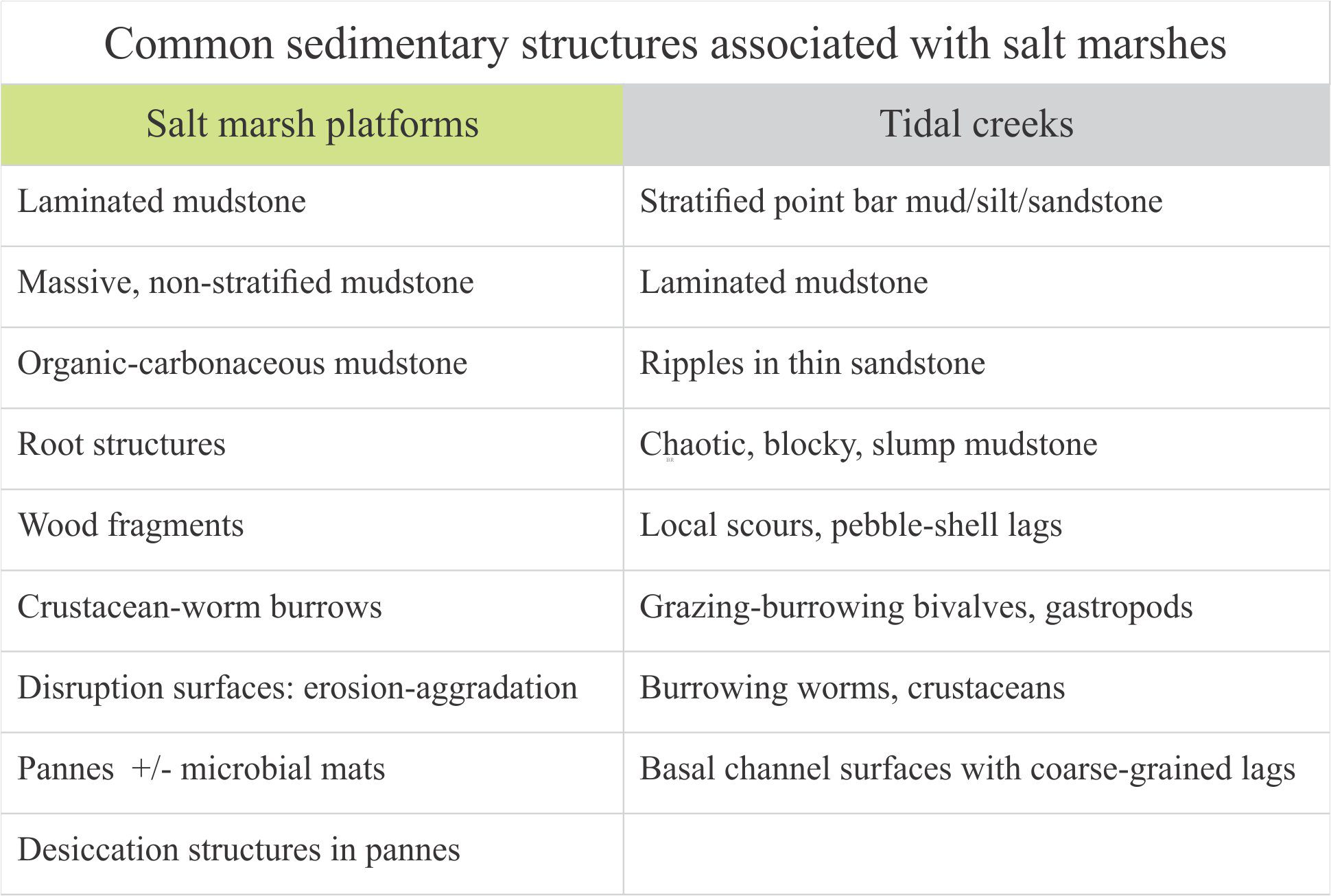
The primacy of beds, bedding, and bedding planes
Beds are the fundamental units of stratigraphy and sedimentology. They are the first things we identify and measure in outcrop, core, and borehole geophysical logs. Beds are the foundations of stratigraphic successions.
Etymologically, the words strata (plural) and stratum (singular) predate the anglicised synonym bed. Leonard da Vinci (1452-1519) and Nicolas Steno (1638-1686) made frequent reference to stratum. The word stratification and its variations are derived from this Latin root. The word bed, in the sense of a resting place or dug plot goes back to Proto-Indo-European roots (about 5000 years back). The context of a sea-bed, where things come to rest, derives from the 16th C Old English bedd. The geological context, as in a stratum, dates from late 17th C although frequent use in scientific literature probably had to wait for James Hutton’s opus (1788), William Smith’s regional geological maps (1819-1824), and Charles Lyell’s ‘Principles’ (1st editions 1830-1833).
Definition of bedding
Beds are sedimentary layers. They usually have observable boundaries top and bottom, referred to as bedding planes. These bounding surfaces are either abrupt where the bedding plane is well defined (you can put your finger on it), or gradational where the compositional or textural change from one bed to the next occurs over some thickness of sediment. Bedding planes demark changes in sediment texture, structure, and/or composition that signify a change in the depositional conditions. The upper bedding plane, if preserved intact, represents a depositional surface – a sediment-air or sediment-water interface. Beds form in all types of sediment: carbonate, siliciclastic, volcaniclastic, chemical. Deposition takes place within a broad spectrum of environmental conditions.
Original orientation
Nicholas Steno (1669) introduced the concept of ‘original horizontality’ where the deposition of sediment at its inception (and therefore bed formation) is approximately horizontal. In reality, the original orientation of a bed will be determined by depositional slope, or paleoslope. This orientation may change during burial compaction, disruption and displacement during soft-sediment deformation, or later tectonism.
Measurable quantities of beds
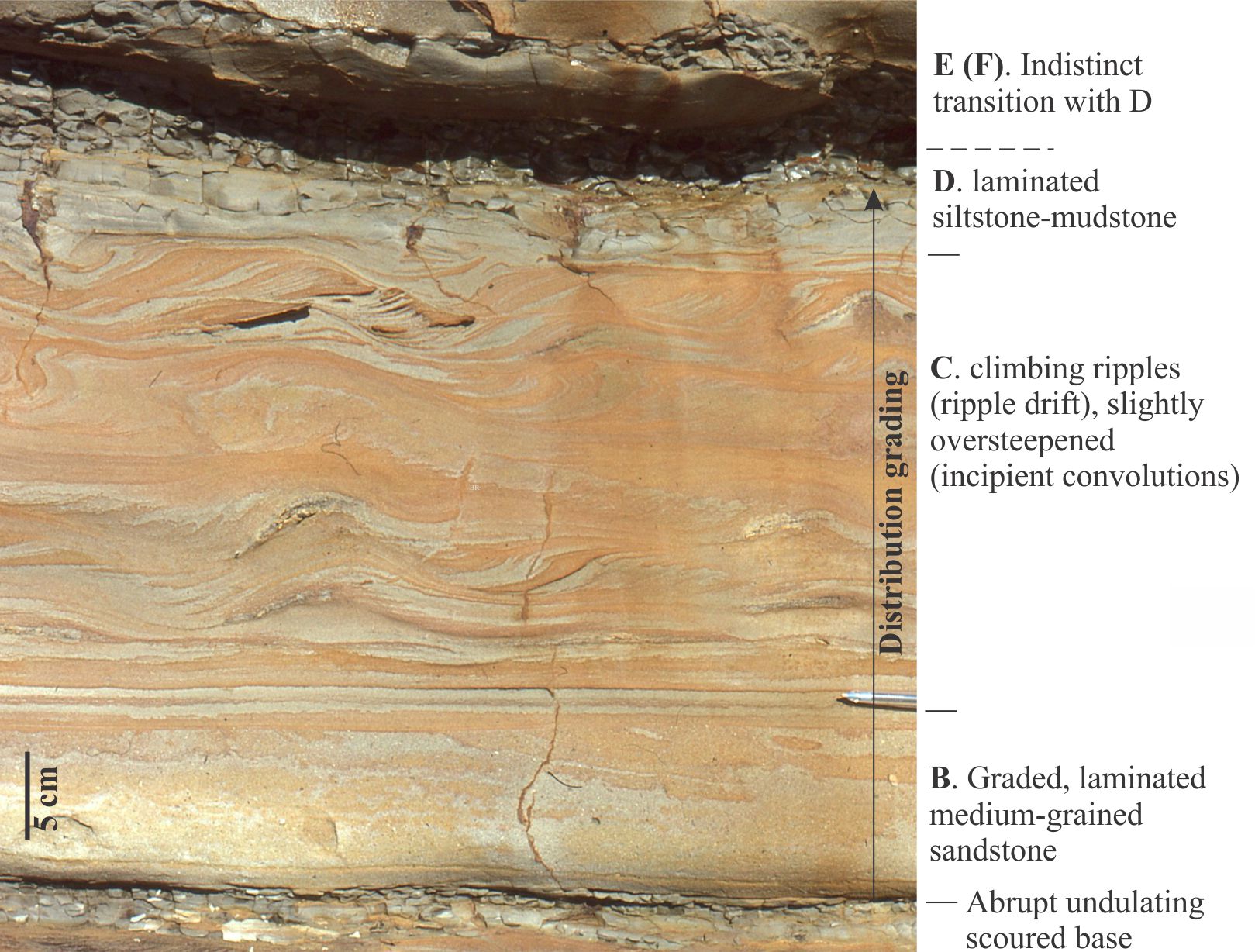
Thickness: Bed thickness is measured between and at right angles to bedding planes. Thickness can vary from millimetres to many 10s of metres depending on the depositional conditions, such as he continuity of sediment supply. For example, slow deposition from suspension in a lake or deep sea can produce millimetre thick laminae, whereas deposition from a debris flow or pyroclastic density current may be metres thick.
Bed geometry: This is usually identified by the 2D and 3D shape of the bedding planes. The scale of observation, particularly in terms of lateral extent, is not codified but is commonly taken to be at least at outcrop scale. As a general rule, bedding is most easily identified at distance from an outcrop – the closer you get, the more complicated it becomes; the point bar deposits shown below illustrate this problem. Common geometric forms include:
- Parallel bedding where bedding planes are parallel at outcrop scale and beyond. Laterally extensive parallel bedding can often be observed in cliff and mountain side exposures. Classic examples occur in flysch-turbidite successions where parallel beds are stacked 100s of metres thick.

Well-developed parallel bedding in an Early Miocene turbidite-debris flow succession. There is a huge range of bed thicknesses here, from 1-2 cm to about 300 cm. Individual beds can be trace laterally for a few hundred metres. Goat Island Marine Reserve, New Zealand.
- Wedge-shaped bedding where bedding planes are not parallel and meet at a pinch out. Theoretically, all beds are probably wedge shaped. At a local scale, examples of this type of bedding include sandstone wedges on a fluvial point bar, and gravel bars in flood-dominated channels on the active portion of an alluvial fan.
- Scour shaped bedding: This type is analogous to wedge-shaped beds, but the lower bounding surface is concave upwards. Examples of this type are commonly attributed to channels and channel-forming processes.

Lateral accretion in this Carboniferous point bar is characterised by discontinuous wedge-shaped sand beds interleaved with siltstone-mudstone layers. From a distance, (left) the inclined lateral accretion beds look quasi-continuous, but their complexity becomes apparent on closer inspection (right). Kentucky, Highway I64.

An amalgamation of sandstone beds filling a fluvial channel – the lowest bed has a distinctive concave-upward bedding plane. Dunvegan Formation (Cretaceous), Alberta.
Internal organization:
- Massive bedding – relatively homogenous and structureless throughout.
- Graded bedding – a change in grain size from bottom to top (e.g., normal, reverse).
- Crossbedded – a variety of bedforms are possible depending on sediment grade and current velocity.
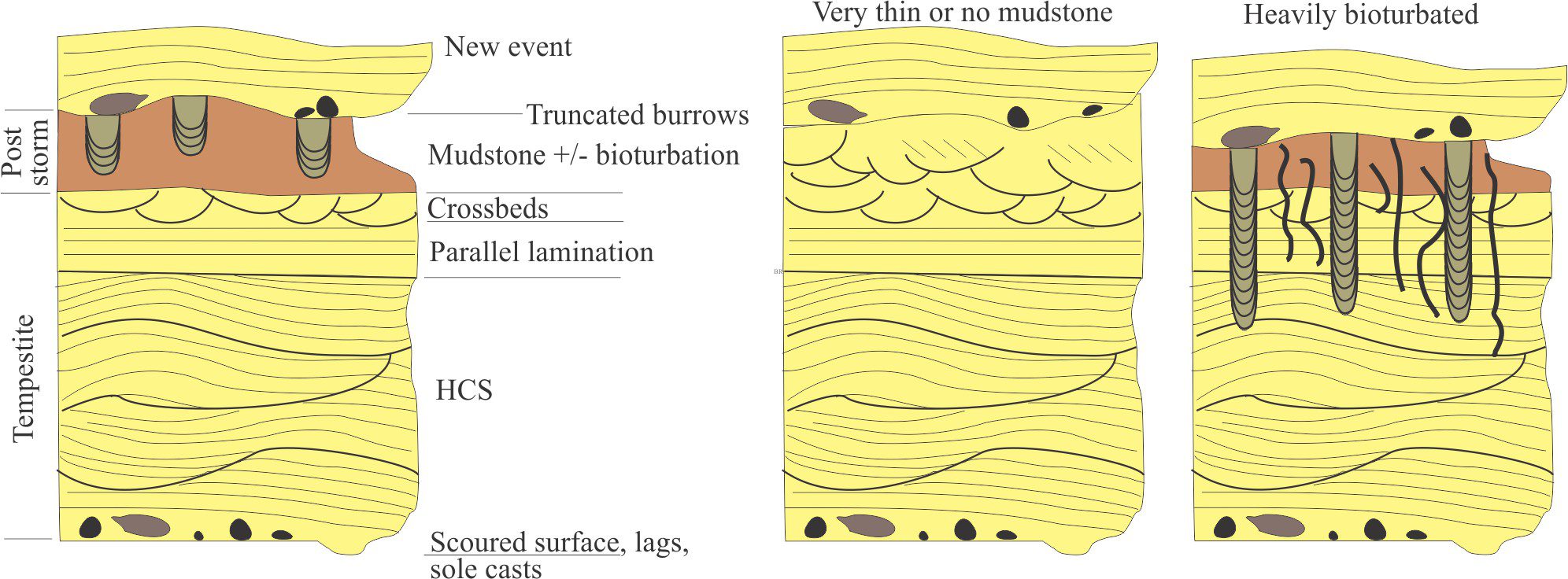
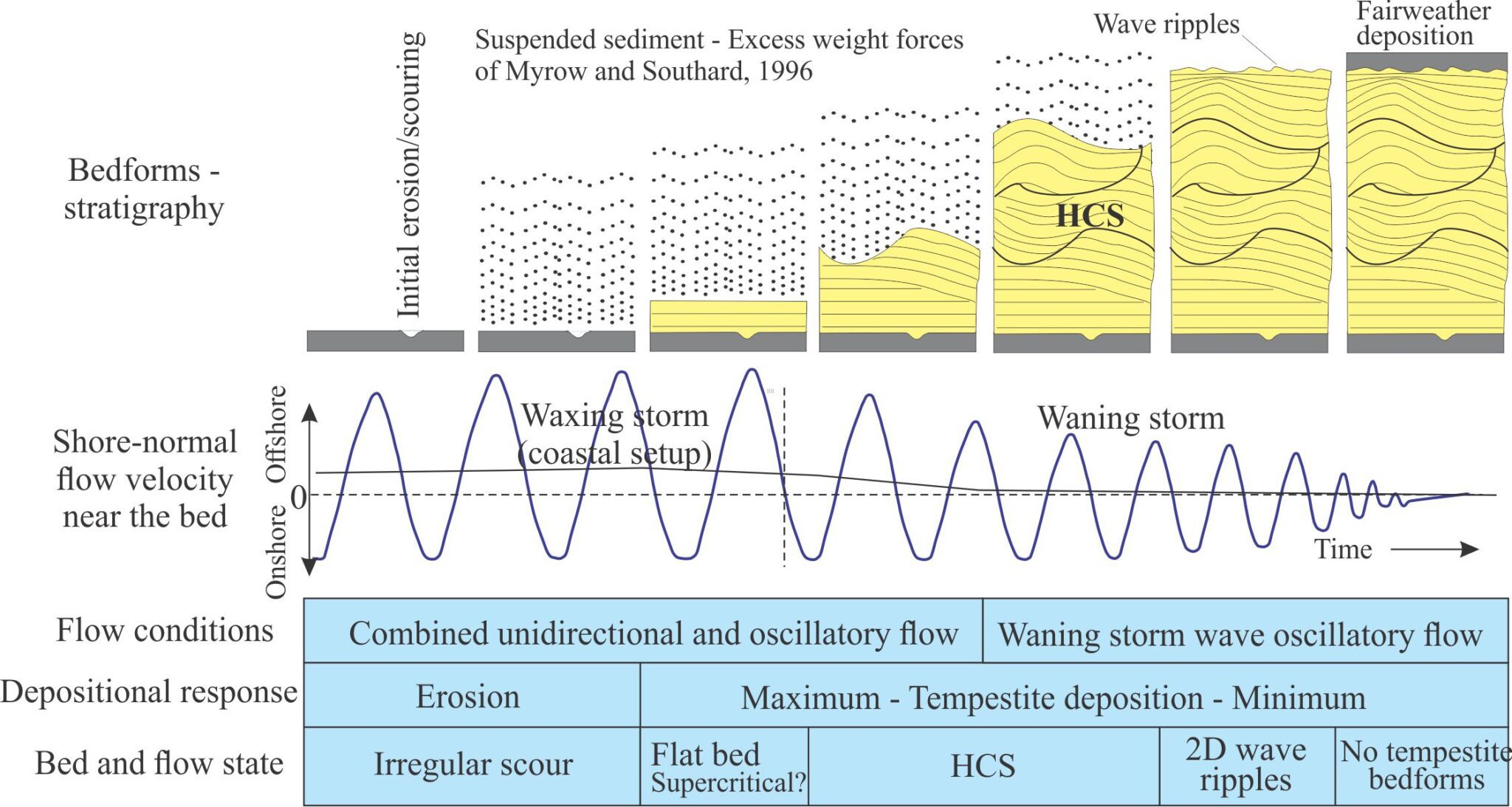
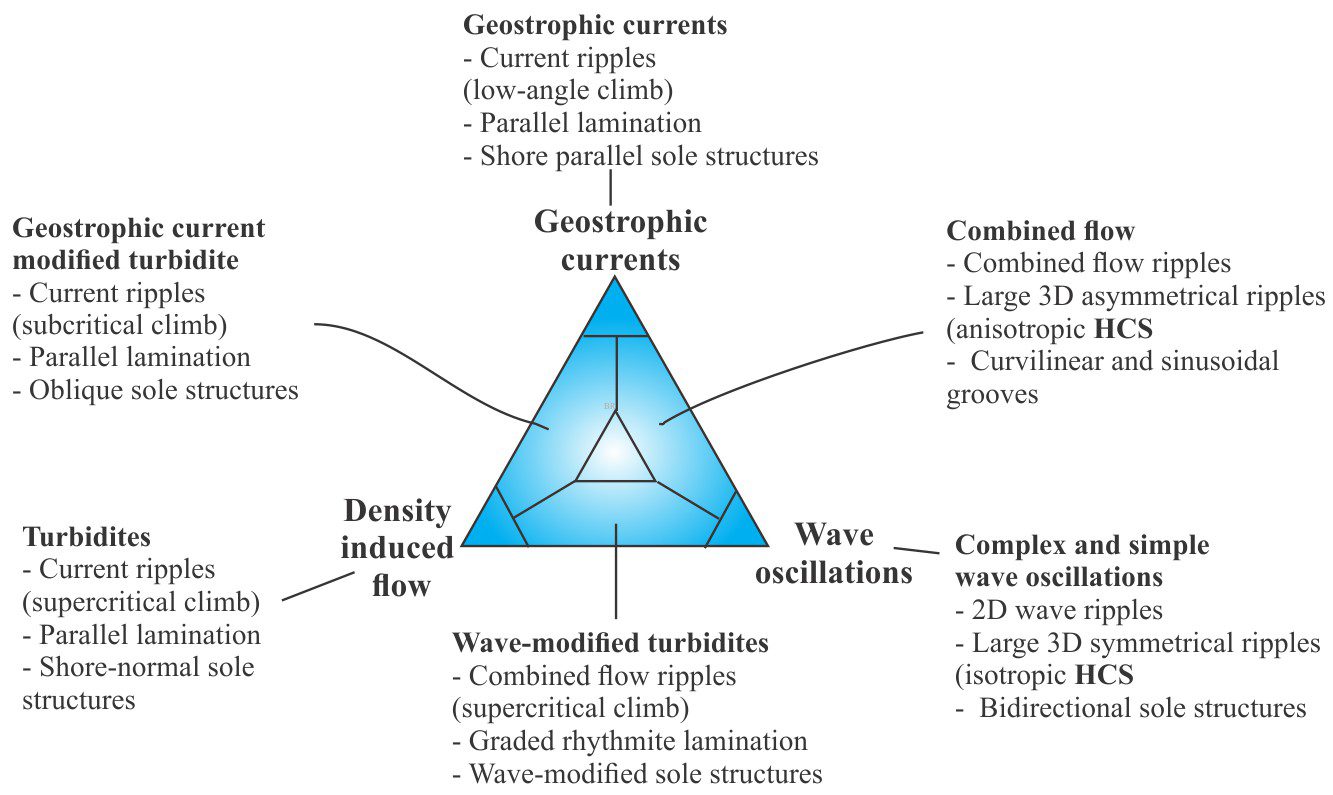
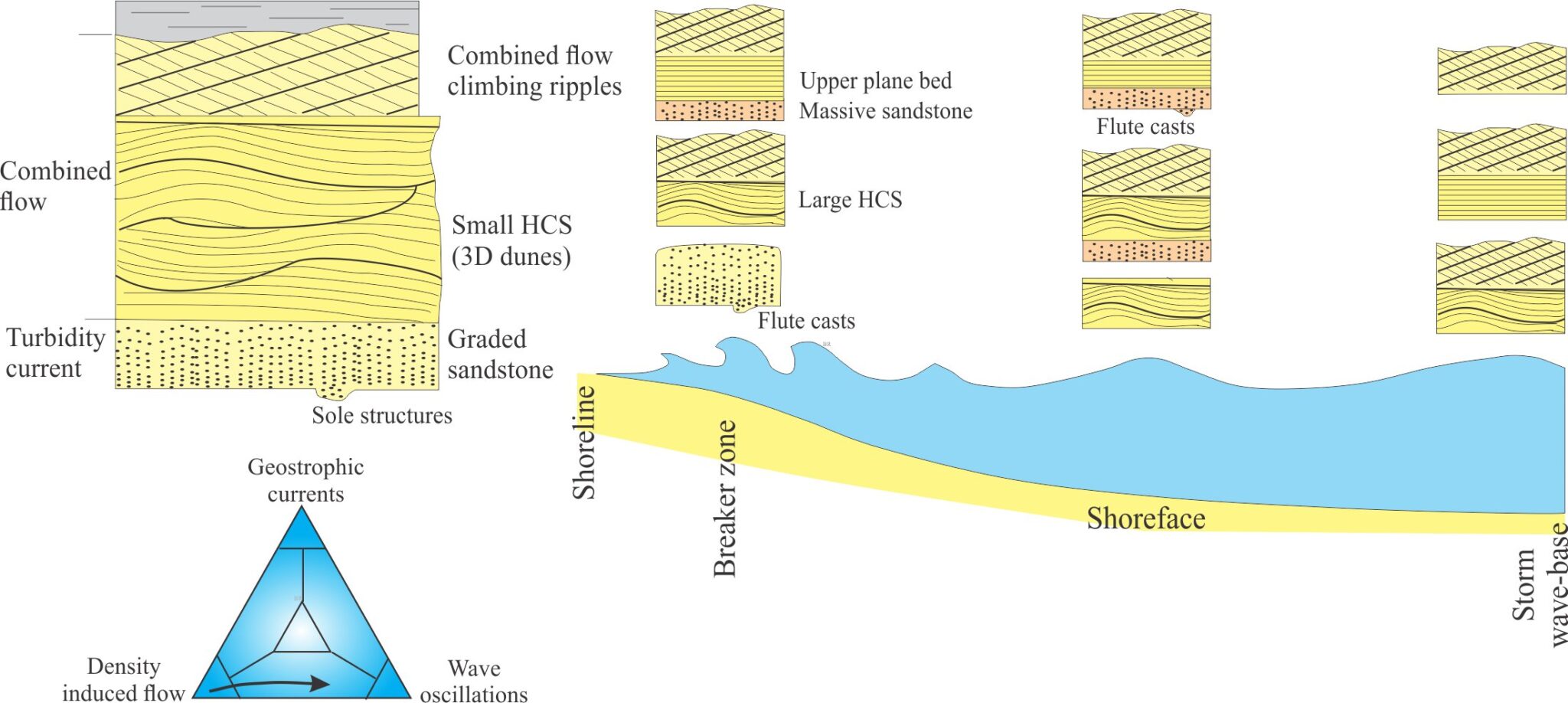
- Event beds: Within any succession, there may be beds that stand out because of an abrupt change in thickness, geometry, or composition – they signify a unique event. For example, a succession of bedded sandstone may be interrupted by a bed containing large mud rip-up clasts, signifying an unusual event such as a storm deposit, or beds that record slumping and soft-sediment deformation, or clasts of different composition that may indicate a change in sediment source (provenance).
- Marker beds: A bit like event beds except they can be traced over large distances across a sedimentary basin. Common examples are volcanic ash beds that represent single eruptions. Minerals in the ash are also potentially useful for radiometric dating the event. Beds like these are important because they approximate chronostratigraphic surfaces and can be used to correlate widely distributed successions.
- Crystal size grading: This applies to chemical sediments. Notable examples include bottom-precipitated evaporite minerals like gypsum and halite, and bedded chert.
- The internal organization of all bed types can be modified by bioturbation, compaction, and post-depositional soft-sediment deformation.

A very distinctive event bed in the Lower Miocene Waitemata Basin, Auckland, consisting of slumped, pulled-apart, folded, and partially liquified sandy turbidite beds. The basal contact is an undeformed glide-plane – a surface over which the entire mass transport deposit moved. The upper bedding plane is irregular, reflecting the relief on top of the slump package, and over which the next sediment gravity flow was deposited.

Thin wavy, undulating and discontinuous beds of gypsum that precipitated at the interface between a salt lake (salar) floor and the overlying brine. Each bed is about 20 mm thick. The gypsum crystals grew vertically from the salar floor. Chilean Altiplano. Probably Late Pleistocene.
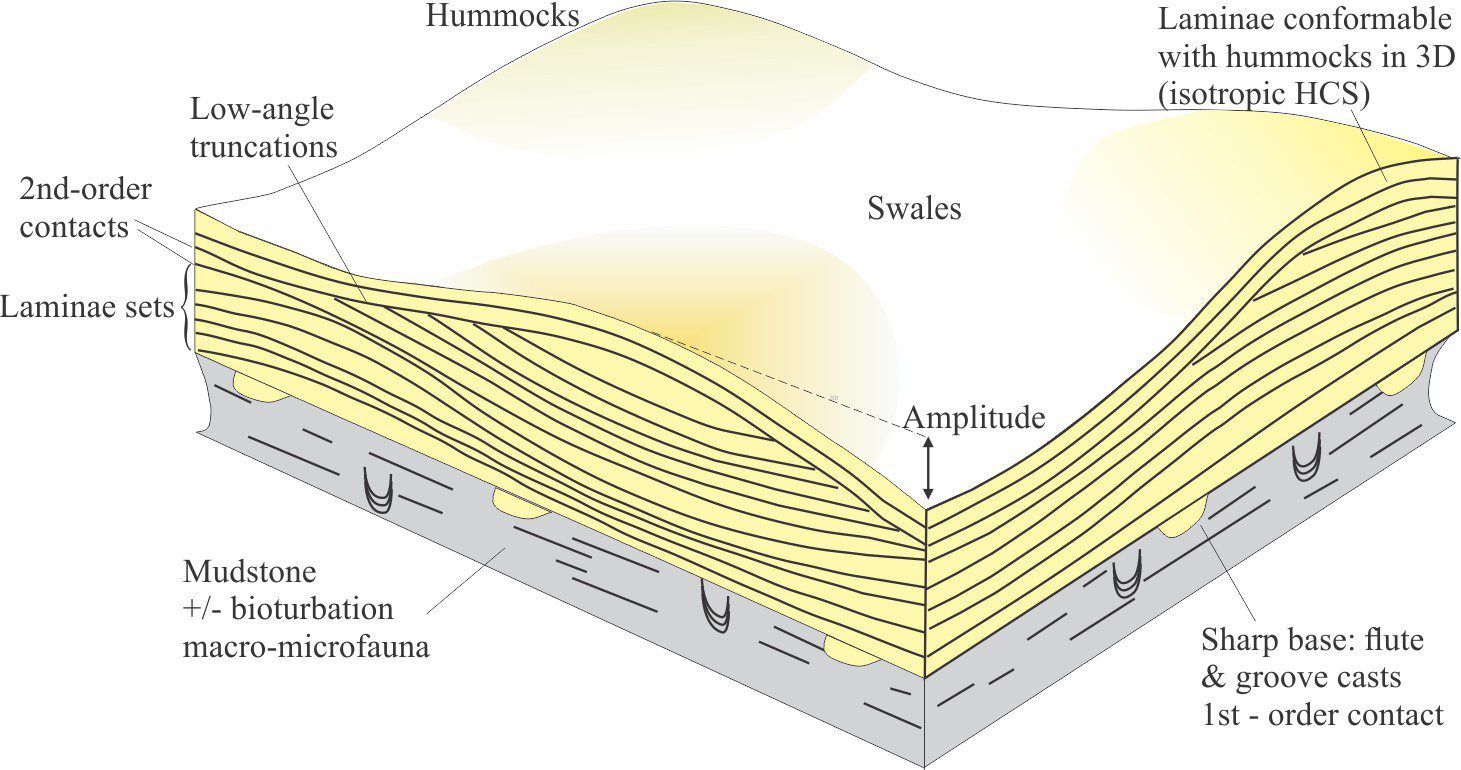
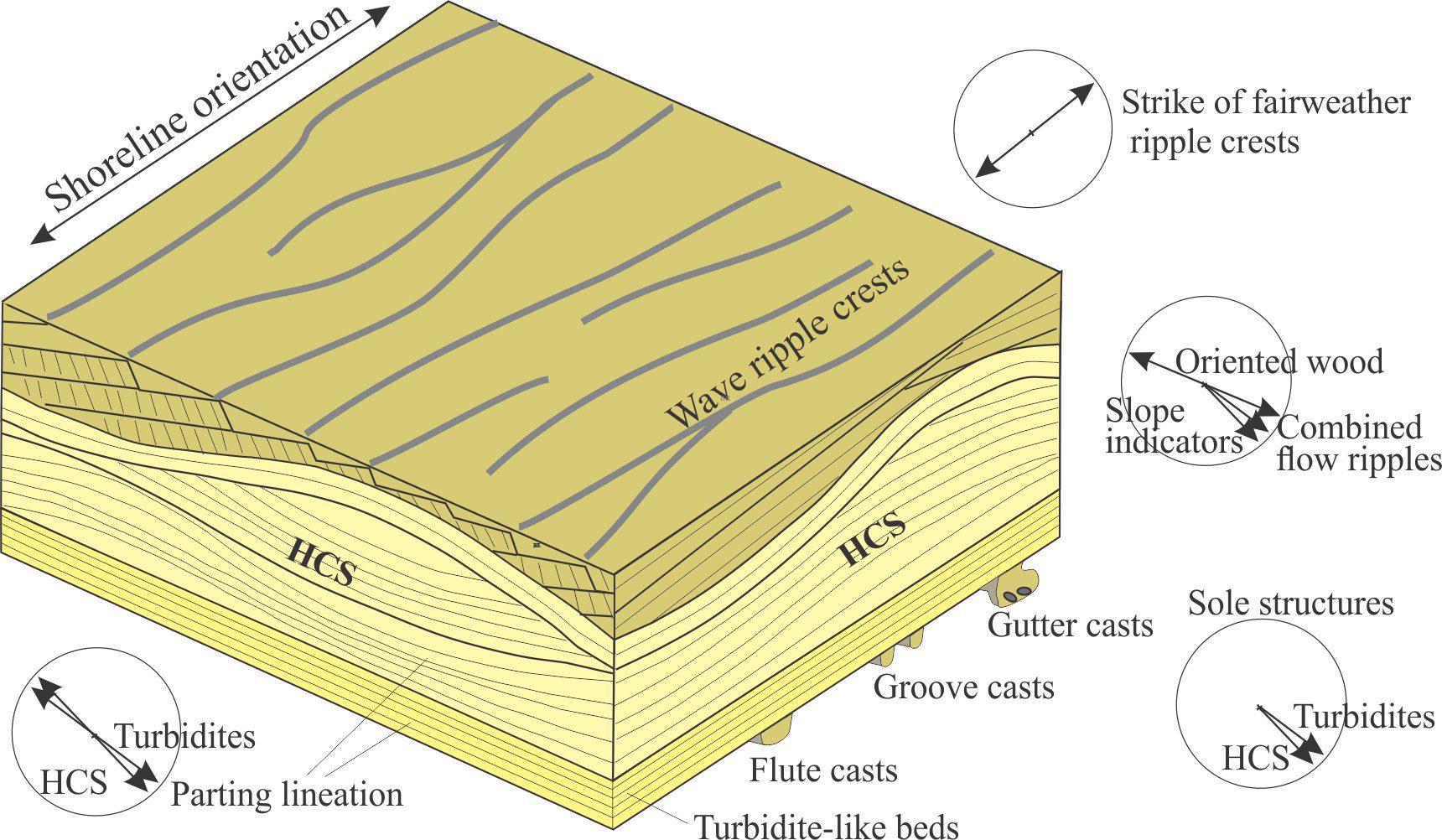
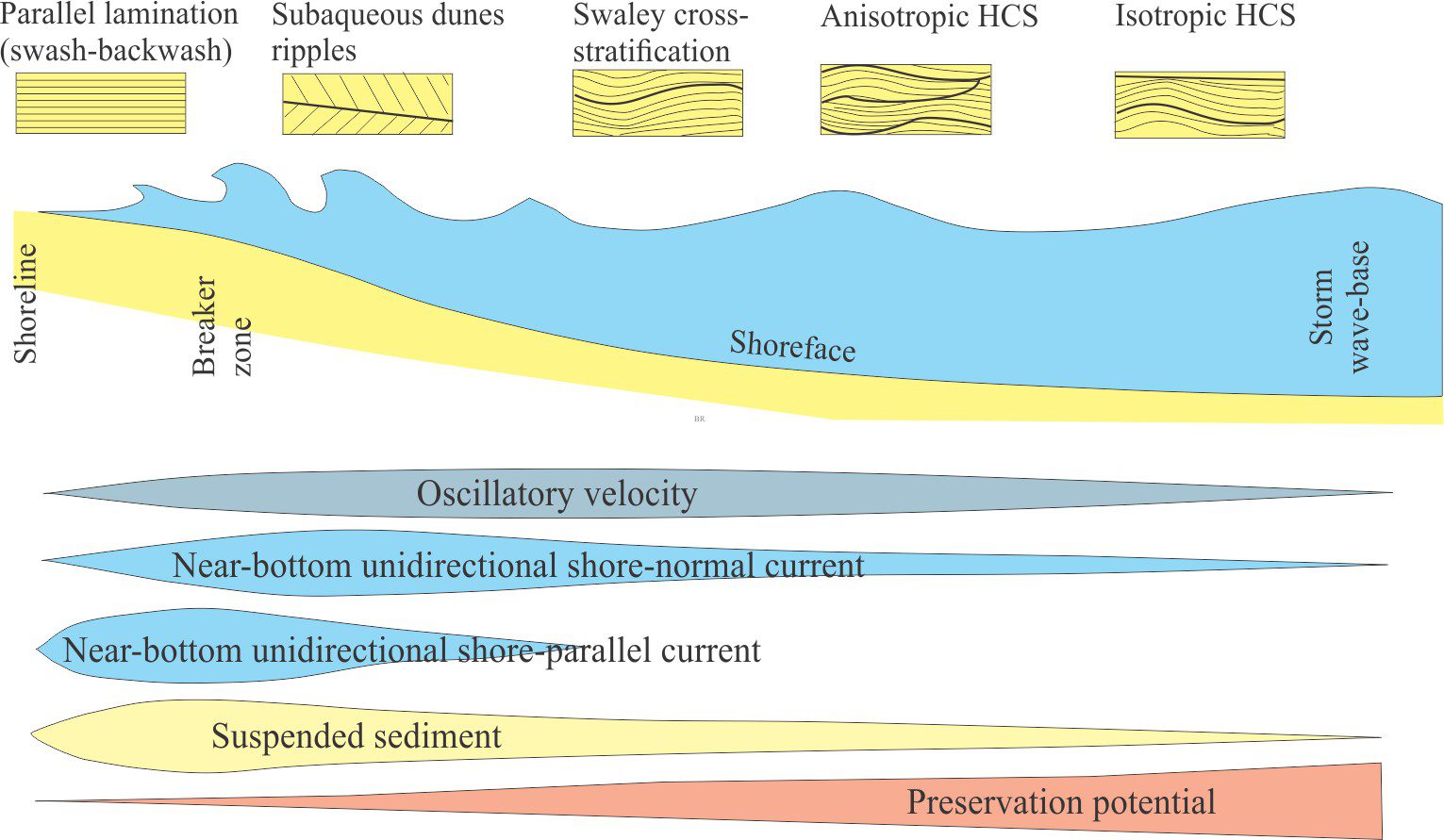
Bedding plane geometry: How a bedding plane is described depends on the resolution of our observations. From a distance, a bedding plane may appear relatively flat or featureless. On closer inspection of the same plane, we might observe undulations that result from large-scale variations in thickness, for example the upper surface of dune bedforms, or large clasts that protrude into the overlying bed; in both cases the departures from ‘flatness’ are produced during the underlying event. However, in many depositional settings, bedding planes are scoured – in this case the scouring is usually associated with the succeeding event. Indeed, erosion and scouring can remove entire beds. Bedding plane irregularities can also result from post-depositional compaction.

The channel-like, lower bounding surface of a crossbedded fluvial sandstone bed has eroded the underlying shelf deposits, in places removing several beds. The fluvial sandstone was deposited during a sea level lowstand when terrestrial drainage extended across the exposed shelf. Jurassic Bowser Basin, northern British Columbia.

Andesite boulders up to 40 cm across protrude through the upper bedding plane of a lahar where they are draped by later, airfall ash and lapilli (white to red-brown layered beds near the top of the exposure). Pliocene Karioi volcano, Raglan, New Zealand.
The significance of bedding planes
- Chronostratigraphic significance of a bed: Every bed represents a period of mechanical or chemical deposition; they are depositional events. The duration of an event can be measured in seconds through millennia. Except under controlled experimental conditions (e.g., flumes), we do not know the duration of these events. We can attempt to find an average duration for a succession, by dividing the number of events (a bed count) by the total time (assuming we can measure the total time represented by the succession), but even this method is woefully inadequate because of…
- Bedding planes as hiatal surfaces: Bedding planes represent the cessation of depositional events. What is unknown is the length of time between the end of one event and the beginning of the next event. A couple of examples: A bed deposited during a river flood is overlain abruptly by a second, similar bed. Was the second bed deposited during a different period of river flooding, or does it represent a very short- duration shift in depositional locus during the same event (maybe the channel axis shifted laterally, or perhaps there was a surge in flow)? In comparison, turbidity currents leave a well-defined and identifiable depositional record, such as the Bouma sequence. Deposition of coarse-grained intervals (A and B) is probably rapid (minutes, hours, days) but the finer-grained parts of the depositional event may take years to complete. The hiatus between this event and the next could well be measured in 100s or 1000s of years.
- Dip and strike: Any description of beds requires us to position them geographically (e.g., latitude-longitude, UTM grids) and to orient them in 3D space. Dip and strike provide unique measures of bed attitude.
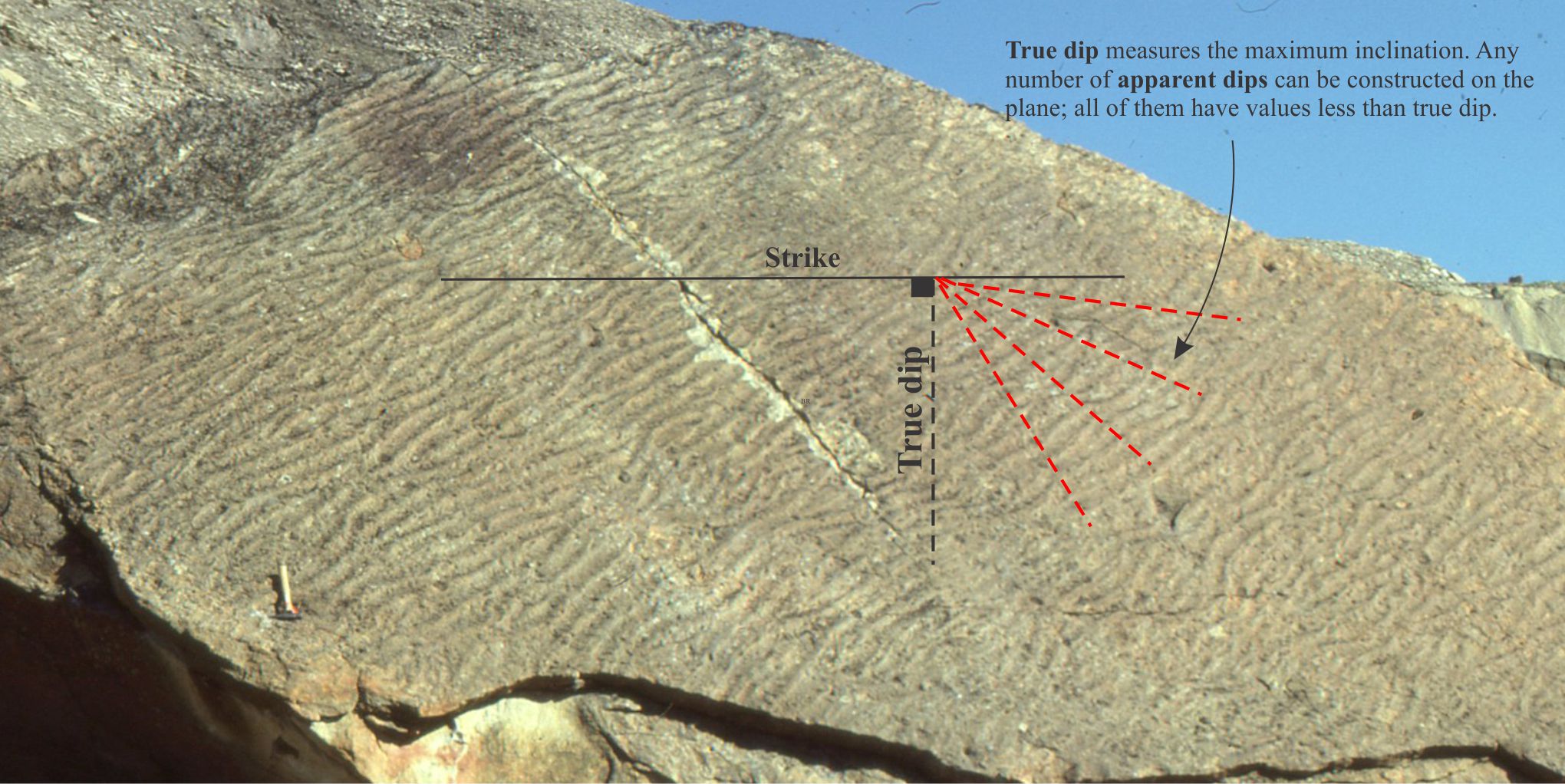
A tilted bedding plane covered with current ripples. The strike, true dip, and apparent dips of are indicated. Paleocene, Ellesmere Island, Arctic Canada.
Things that are not beds
- Metamorphic layering: Original bedding can be preserved in low grade metasedimentary and metavolcanic rocks (e.g. subgreenschist to low greenschist grade). High grade rocks (upper greenschist, amphibolite) commonly present compositional layering and through-going foliation that result from alignment of recrystallized sheet silicates like muscovite and biotite. In most cases, original sedimentary bedding has been obliterated.
- Igneous dykes (dikes) and sills: Both represent intrusion of igneous melts into an existing pile of rock. They are not beds.
- Sedimentary dykes: These too are intrusive bodies (of fluid sediment) that are insinuated into an existing pile of sedimentary beds.
- Stratiform iron pans: Bands of nodular limonite and goethite commonly precipitate within existing beds of porous sediment, in response to groundwater infiltration and watertable fluctuations. The iron bands commonly mimic bedding because of the permeability advantage, but can also cross-cut bedding.

Discontinuous limonite iron pans superficially mimic the left-dipping bedding in these Pleistocene sand dune deposits, but the pans also crosscut the dune bedding (arrows). The iron pans post-date dune deposition and formed as subsurface precipitates of iron oxide during older, fluctuating watertables. Kariotahi, New Zealand.
Other posts that introduce basic methods of rock description, mapping, and structural analysis
Solving the three-point problem
The Rule of Vs in geological mapping
Plotting a structural contour map
Stereographic projection – the basics
Stereographic projection of linear measurements
Stereographic projection – unfolding folds
Stereographic projection – poles to planes
Faults – some common terminology
Thrust faults: Some common terminology
Strike-slip faults: Some terminology
Using S and Z folds to decipher large-scale structures